有機性廃棄物の有効利用の一つにメタン発酵がある。メタン発酵は、酸発酵細菌にて有機物をいったんギ酸、酢酸、乳酸などに変換させ、それをメタン菌に利用させる。60℃付近にて有機物から酸を生成する好熱性細菌を探索していたときに、偶然に細菌鉱物化現象は発見された。
単離した細菌は中等度好熱性細菌
単離した細菌は、胞子形成能力のあるグラム陽性細菌で 長径2-5μmの桿菌であった(Fig 1A, B)。胞子は栄養細胞の端に形成される(Fig. 1C, D)。自家蛍光があり365nmの光を照射すると、青く発色する(Fig. 1E, F)。SCD寒天培地(日水製薬)において、50〜70℃の範囲で生育が認められた。生育最適温度は60℃であるので、中等度好熱性細菌であるといえる。生育可能なpHは狭くpH7-8であった。16SrDNAの塩基配列1484 bpを決定し、BLASTによるホモロジー検索を行った。その結果Geobacillus thermoglucosidasius ATCC43742のそれともっとも相同性が高かったことより、単離細菌をGeobacillus thermoglucosidasius NY05と命名した。Geobacillus thermoglucosidasius NY05は理化学研究所微生物材料開発室のタイプカルチャーとして保存されている(JCM31848) 。
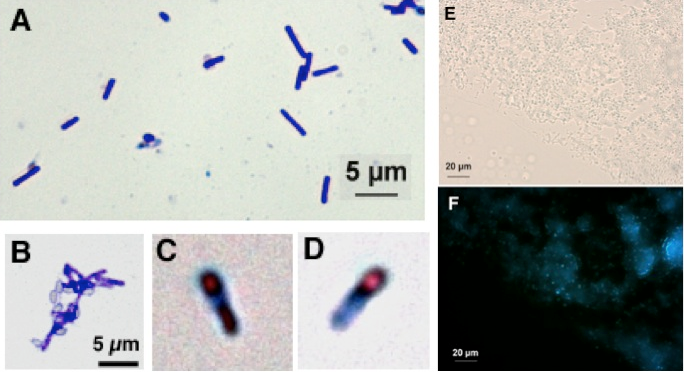
Fig. 1 Geobacillus thermoglucosidasius NY05の光学顕微鏡写真
低級脂肪酸を鉱物に変換する
本研究の新規性は、好熱性細菌が鉱物の単結晶を形成することを発見した点と、単結晶を形成させる新手法を開発した点にある。新手法は細胞を地球にあり得ない環境に曝すことになるので、特異状態であるといえる。
SCD寒天培地にG. thermoglucosidasius NY05を植え、60℃、18-24時間培保温すると、新鮮なバイオマスが得られる。このバイオマスを25 mM酢酸ナトリウムと7 mM塩化カルシウムを含む1.5%寒天ゲル(結晶形成ゲル)表面に付着させ(parent colony)、常圧60℃にて1-4日反応させる。
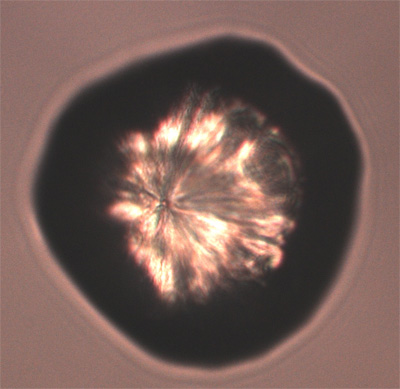
反応後はFig. 2に示すように、parent colonyの中にも外にも鉱物の単結晶が形成されているのがわかる。結晶の大きさは110-130 µmで、形はダンベル型、球形、六方晶型 (Fig. 3)など様々である。この単結晶は蛍光特性があり365±5、480±20、545±15 nmの波長で励起させるとそれぞれ青、緑、赤色の発色が観察された。最大励起波長は369.6 nmで、最大放出波長は446.9 nmであった (Fig. 4)。
G. thermoglucosidasius NY05が鉱物形成に利用できる脂肪酸はC4化合物までで、ギ酸、酢酸、プロピオン酸、酪酸塩を利用して単結晶を形成することが確かめられている。
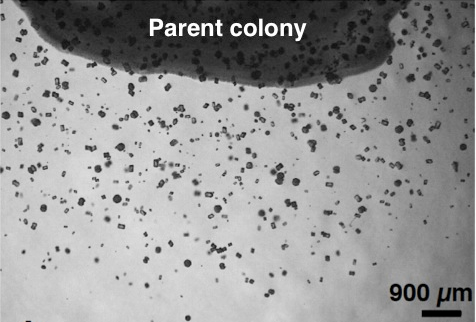

Fig. 2 結晶形成ゲル上のparent colonyと Fig. 3 鉱物結晶の拡大写真
形成された鉱物結晶
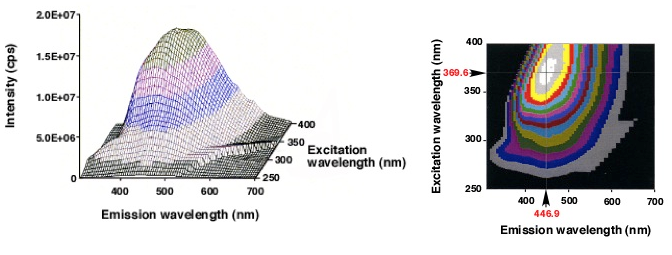
Fig. 4 単結晶の蛍光特性
単結晶の構成元素
エネルギー分散型X線解析により単結晶の元素分析を行ったところ、得られたスペクトルは炭酸カルシウムのそれとよく似ていた。炭素と酸素を除いた元素の構成割合を調べると、炭酸カルシウムはカルシウム100 (atm%)にて構成されているのに対し、単結晶はカルシウム93.4、マグネシウム6.6 (atm%)にて構成されていた。粉末X線回折装置にて分析すると、炭酸カルシウムと単結晶のピーク位置は酷似しており、得られたスペクトルをThe Powder Diffraction Fileのデータベースに照合すると、単結晶は炭酸塩鉱物の中のマグネシウムカルサイトであることが判明した。
酢酸にはカルボキシル炭素(一位)とメチル炭素(二位)がある。一位の炭素を安定同位体13Cでラベルした酢酸を用いてマグネシウムカルサイトを形成させ、その炭素同位体比を調べると、12Cが56.9%であるのに対し、13Cは43.1%であった。二位の炭素を13Cでラベルした酢酸を用いてマグネシウムカルサイトを形成させた場合、その炭素同位体比はそれぞれ62.1、37.9%であった (Fig. 5)。このことはマグネシウムカルサイトを構成する炭素は酢酸由来であることを示している。酢酸分子の一、二位の炭素両者共にマグネシウムカルサイト形成に利用される。特に一位の炭素の方が利用されやすいといえる。13Cの比率を両実験で合計すると81%になる。したがってマグネシウムカルサイト炭素の19%は空気中の二酸化炭素由来であると予想される。
G. thermoglucosidasius NY05が仲介する化学反応によって形成される単結晶にはマグネシウムが6.6% (atm%)含まれている。結晶形成ゲルにはマグネシウムは含まれないので、単結晶を構成するマグネシウムはG. thermoglucosidasius NY05の栄養細胞中に存在するマグネシウム由来であると考えられる。SCD培地にて培養されたG. thermoglucosidasius の栄養細胞中に含まれるマグネシウム量を調べてみると、乾燥重量1mg当たり2.2 µg含まれていた。この値はEnterococcus duransやBacillus subtilisのそれと比較すると高かった。
G. thermoglucosidasiusの栄養細胞内のカルシウム含有量を、マグネシウムカルサイトを形成する能力が認めなれていない他の中等度好熱性細菌のそれと比較したところ、G. thermoglucosidasius NY05のカルシウム含有量は湿重量35mg中1.75mg含んでおり、他の中等度好熱性細菌よりも4-5倍高かった。
結晶形成ゲル中にポリアルクリル酸、EDTAを含ませておくと、濃度が増すに従って、parent colony中に形成されるマグネシウムカルサイトの結晶数は減少した。それぞれ0.6 mg/ml、5 mMはマグネシウムカルサイト形成を完全に阻害した。ポリアクリル酸はカルシウムと親和性が高い物質であり、EDTAは金属キレート剤である。これらの物質は反応場のカルシウムを捕獲し、結晶形成の進行を阻害しているといえる。マグネシウムカルサイト形成には、結晶形成ゲル中に含まれるカルシウムが利用されることは明白である。
N,N-dicyclohexylcarbodiimide(DCCD)はH+-ATPase(エネルギー転移)阻害剤またはV-ATPase阻害剤として知られている。DCCDを結晶形成ゲル中に添加すると結晶数は減少することから、G. thermoglucosidasiusの栄養細胞によるカルシウムの取り込みはATP依存的ではないかと考えられる。
G. thermoglucosidasius NY05は、他の細菌よりは細胞内にマグネシウムやカルシウムを多く蓄える性質がある。マグネシウムやカルシウムを細胞内に取込むチャネルタンパク質を高度に発達させているのではないかと予想される。マグネシウムおよびカルシウムの細胞内への取込みはマグネシウムカルサイト形成には重要な役割をもつと思われる。
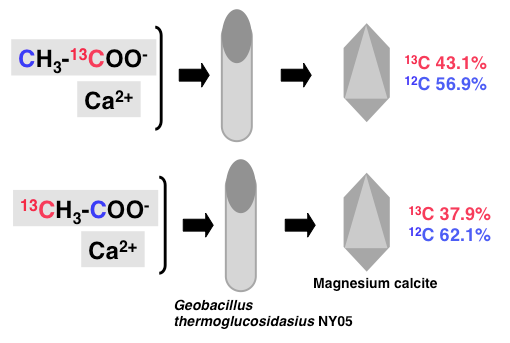
Fig. 5 安定同位体13Cでラベルした酢酸を用いて形成されたカルサイトの安定同位体の割合
G. thermoglucosidasius NY05の結晶形成ゲル上での運動性
直径9cmのペトリディッシュに結晶形成ゲルを固化させ、中央部にG. thermoglucosidasius NY05のバイオマスを置き、常圧60℃に保った。24時間以降結晶形成ゲルを切取り、SCD寒天培地に貼付け、適温に置くと、コロニーが形成されるので、栄養細胞が移動した位置がわかる。コロニーが形成される位置は次第にparent colonyから離れた場所に確認されるようになり、栄養細胞はparent colonyの中心から最大30mmまで遊泳することがわかった。マグネシウムカルサイト単結晶の出現数も168時間までに次第にparent colonyの中心から外側に向けて増加している (Fig. 6)。
結晶形成ゲルのゲル濃度と、parent colonyの縁からのカルサイトの形成距離の関係を調べると、寒天ゲル1.0~1.5%では7 mmまでカルサイト単結晶が形成されるが、2、 3%ではそれぞれ4、2mmと次第に狭まってくる。4~5%ではparent colonyの外側にはまったくカルサイトは形成されなかった (Fig. 7)。ゲル濃度が高まるとカルサイト形成距離が狭まるのは、栄養細胞の運動性が弱まっている証拠である。ゲル濃度が高いと抵抗が大きくなり、運動性が制限されることを示している。以上のことよりparent colonyの外側にもマグネシウムカルサイトが形成されるのは、parent colonyから泳ぎ出ている栄養細胞が元になっていると考えられる。
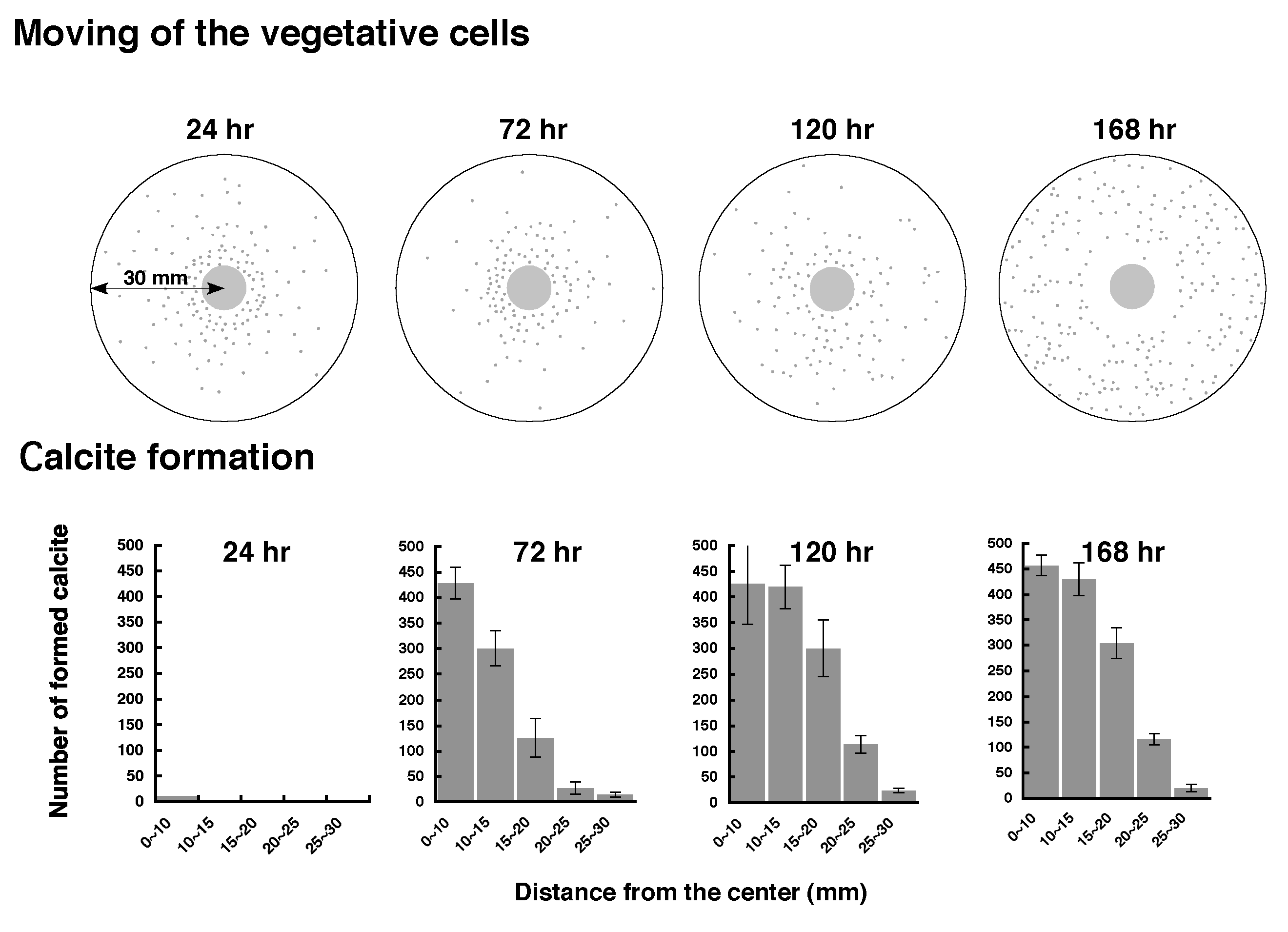

Fig. 6 結晶形成ゲル上を活発に遊泳するG. thermoglucosidasius NY05と Fig. 7 結晶形成ゲルの各ゲル濃度 (1.0-5.0%)におけるカルサイト形成域 parent colony中心からカルサイトとの距離およびその形成数
カルサイトに閉じ込められた生命
カルサイトを実体顕微鏡にて注意深く観察すると、中心部に結晶成長の核を見出すことができる(Fig. 8A)。カルサイト単結晶をSCD寒天培地に置き、60℃にて保温すると、そこから細菌のコロニーが生育してくる (Fig.8E)。顕微鏡にて観察すると、カルサイト単結晶の中央部が割れ、光の透過度が増している様子が確認できる(Fig. 8B)。走査型電子顕微鏡による観察でも、割れたカルサイトの中心部には核らしき粒子が見えるし (Fig. 8C)、結晶中央部に明確な亀裂を生じたダンベル型結晶も確認することができる (Fig. 8D)。
カルサイト単結晶を1N塩酸にて溶解させると、マラカイトグリーンにて染色される300-400個の粒子が残存する。この粒子はその大きさや形も加味すれば、G. thermoglucosidasius NY05の内生胞子である可能性が高い。内生胞子は300-400個集まり、カルサイト結晶成長の核になると考えている。この粒子からはDNAが検出され、また分子量75kDaのタンパク質も検出された。MALDI-TOF MASS spectrometryを用いて質量分析し、得られた分析データを既存のデータベースと照合すると、このタンパク質はBacillus subtilis由来のN-acetyltransferase YitHに最も類似していた。
グルコースはG. thermoglucosidasius NY05の胞子形成を強く阻害する。グルコース6 mMを含む結晶形成ゲルを用いた場合、カルサイト形成は完全に阻害された。グルコースがカルサイト形成を阻害することは、カルサイトの結晶成長には胞子が不可欠であることを支持する。
栄養的に好条件になればカルサイト結晶に亀裂が入り、栄養細胞のコロニーを放出するということは、カルサイトは生命を内包した鉱物であるといえる。
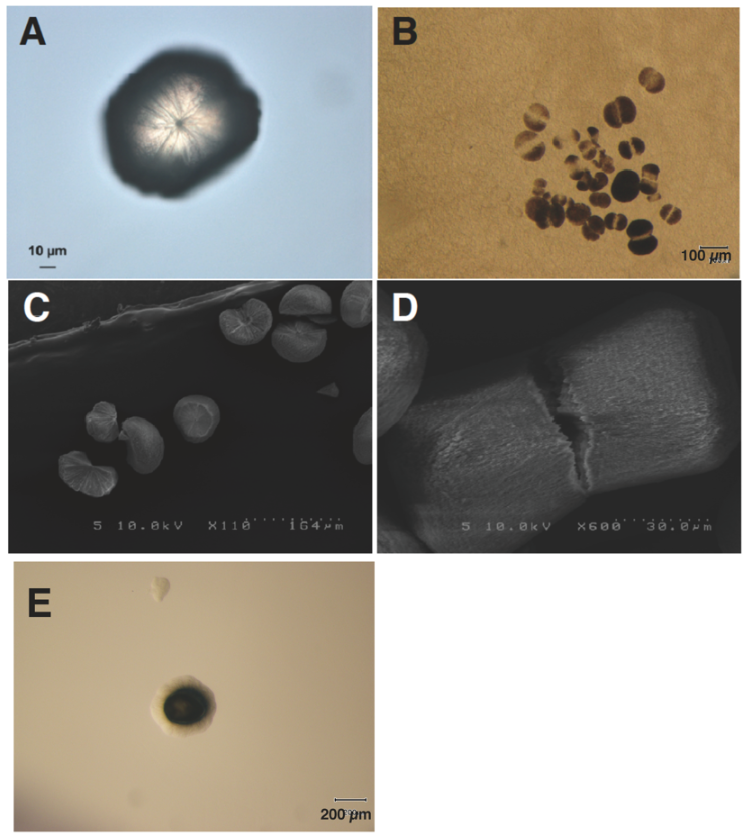
Fig. 8 カルサイト結晶の核および亀裂と成長するG. thermoglucosidasius NY05のコロニー
希土類やレアメタルを用いない蛍光体の調製
炭酸カルシウム蛍光体の調製には、Euなどの希土類やレアメタルを蛍光賦活剤として母材へ固溶させなければならない。母材の構造中に賦活剤を置換固溶させるためには高温処理する必要があるため、製造コストが上昇する。G. thermoglucosidasius NY05の触媒作用によって形成される鉱物結晶は、希土類やレアメタルを使用しないカルサイト蛍光体である。様々な重金属イオンをドープした結晶形成ゲルを用いて鉱物結晶を形成させ、それらの蛍光特性と結晶の形態変化を調べた。カルサイト結晶中に置換固溶するのはマグネシウム、リン、マンガン、ストロンチウムであった。2 mMマグネシウムの結晶形成ゲルへのドーブはカルサイトのマグネシウム置換固溶量を21-22%に増加させた。なかんずく2 mM アルミニウム、ケイ素、リンの結晶形成ゲルへのドープは、カルサイトの蛍光強度を1.7倍 (EX 360 nm, EM 465 nm) に増加させた。反対にマンガン、ストロンチウム、鉄、コバルトは蛍光強度を減少させた。マグネシウム、リンをドープすると、それぞれ表面が滑らかで球形の結晶型、薄板状の結晶型へと変化した。蛍光顕微鏡による観察では、カルサイト結晶の青色、緑色、赤色の蛍光を確認した。高価な希土類やレアメタルを使用せず、低コストで環境負荷の少ない蛍光体を開発できる道筋がつけられた。
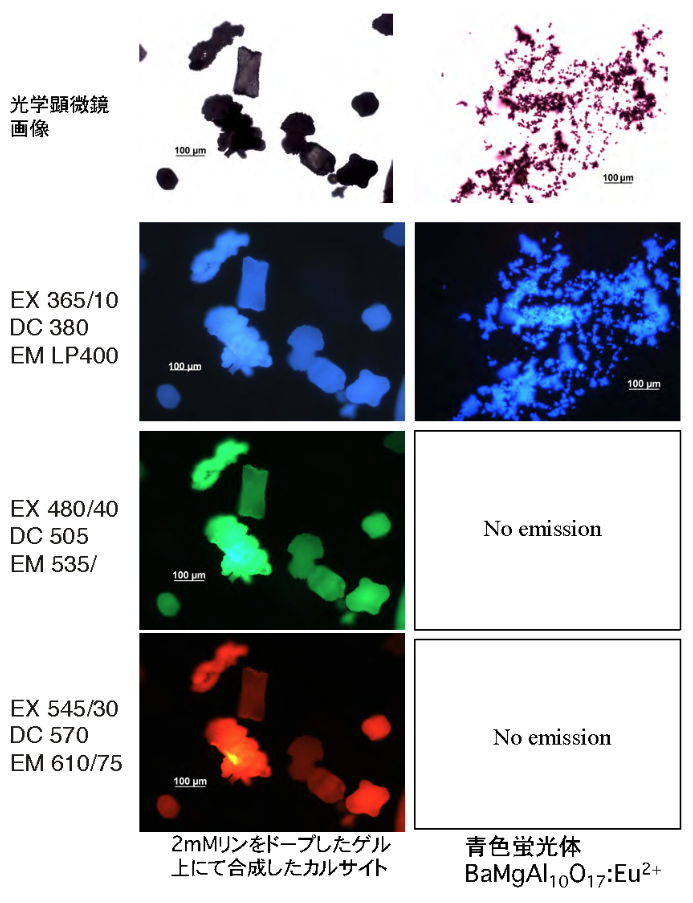
Fig. 9 蛍光強度の増したカルサイトと青色蛍光体の比較
パンスペルミア説支持へ向けての取組み
(作成中・・・)